

What Do Ribosomes Do In An Animal Cell
| Cell biology | |
|---|---|
| Animal prison cell diagram | |
 Components of a typical animal prison cell:
|

Figure 1: Ribosomes get together polymeric protein molecules whose sequence is controlled by the sequence of messenger RNA molecules. This is required by all living cells and associated viruses.
Ribosomes ( ), also chosen Palade granules (afterwards discoverer George Palade and due to their granular structure), are macromolecular machines, found inside all cells, that perform biological poly peptide synthesis (mRNA translation). Ribosomes link amino acids together in the guild specified past the codons of messenger RNA (mRNA) molecules to form polypeptide chains. Ribosomes consist of two major components: the small and large ribosomal subunits. Each subunit consists of one or more ribosomal RNA (rRNA) molecules and many ribosomal proteins (RPs or r-proteins).[1] [ii] [3] The ribosomes and associated molecules are also known as the translational apparatus.
Overview [edit]
The sequence of Deoxyribonucleic acid that encodes the sequence of the amino acids in a protein is transcribed into a messenger RNA chain. Ribosomes bind to messenger RNAs and use their sequences for determining the correct sequence of amino acids to generate a given protein. Amino acids are selected and carried to the ribosome by transfer RNA (tRNA) molecules, which enter the ribosome and bind to the messenger RNA concatenation via an anti-codon stem loop. For each coding triplet (codon) in the messenger RNA, at that place is a unique transfer RNA that must have the exact anti-codon match, and carries the right amino acid for incorporating into a growing polypeptide chain. Once the poly peptide is produced, it can and then fold to produce a functional three-dimensional structure.
A ribosome is made from complexes of RNAs and proteins and is therefore a ribonucleoprotein complex. Each ribosome is equanimous of small-scale (30S) and large (50S) components, chosen subunits, which are bound to each other:
- (30S) has mainly a decoding function and is as well bound to the mRNA
- (50S) has mainly a catalytic function and is also jump to the aminoacylated tRNAs.
The synthesis of proteins from their building blocks takes place in four phases: initiation, elongation, termination, and recycling. The start codon in all mRNA molecules has the sequence AUG. The stop codon is one of UAA, UAG, or UGA; since at that place are no tRNA molecules that recognize these codons, the ribosome recognizes that translation is complete.[4] When a ribosome finishes reading an mRNA molecule, the two subunits separate and are ordinarily broken up just tin be re-used. Ribosomes are ribozymes, because the catalytic peptidyl transferase activity that links amino acids together is performed by the ribosomal RNA.[five]
Ribosomes are oftentimes associated with the intracellular membranes that make up the rough endoplasmic reticulum.
Ribosomes from leaner, archaea and eukaryotes in the three-domain system resemble each other to a remarkable degree, evidence of a common origin. They differ in their size, sequence, construction, and the ratio of protein to RNA. The differences in structure allow some antibiotics to impale bacteria by inhibiting their ribosomes, while leaving human ribosomes unaffected. In all species, more than i ribosome may move along a single mRNA chain at one time (as a polysome), each "reading" a specific sequence and producing a corresponding protein molecule.
The mitochondrial ribosomes of eukaryotic cells functionally resemble many features of those in bacteria, reflecting the likely evolutionary origin of mitochondria.[half-dozen] [vii]
Discovery [edit]
Ribosomes were first observed in the mid-1950s by Romanian-American prison cell biologist George Emil Palade, using an electron microscope, every bit dense particles or granules.[eight] The term "ribosome" was proposed by scientist Haguenau in the stop of 1958:
During the course of the symposium a semantic difficulty became apparent. To some of the participants, "microsomes" mean the ribonucleoprotein particles of the microsome fraction contaminated by other protein and lipid material; to others, the microsomes consist of protein and lipid contaminated by particles. The phrase "microsomal particles" does not seem acceptable, and "ribonucleoprotein particles of the microsome fraction" is much too awkward. During the meeting, the discussion "ribosome" was suggested, which has a very satisfactory name and a pleasant audio. The present confusion would be eliminated if "ribosome" were adopted to designate ribonucleoprotein particles in sizes ranging from 35 to 100S.
—Albert, Microsomal Particles and Protein Synthesis[9]
Albert Claude, Christian de Duve, and George Emil Palade were jointly awarded the Nobel Prize in Physiology or Medicine, in 1974, for the discovery of the ribosome.[ten] The Nobel Prize in Chemical science 2009 was awarded to Venkatraman Ramakrishnan, Thomas A. Steitz and Ada Due east. Yonath for determining the detailed structure and mechanism of the ribosome.[11]
Structure [edit]

Ribosome rRNA composition for prokaryotic and eukaryotic rRNA
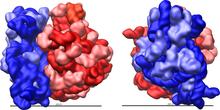
Effigy 2: Large (red) and small (blue) subunit fit together.
The ribosome is a complex cellular machine. It is largely made up of specialized RNA known as ribosomal RNA (rRNA) also as dozens of distinct proteins (the exact number varies slightly between species). The ribosomal proteins and rRNAs are arranged into two distinct ribosomal pieces of different sizes, known by and large as the large and pocket-sized subunit of the ribosome. Ribosomes consist of ii subunits that fit together (Figure 2) and work every bit i to translate the mRNA into a polypeptide chain during protein synthesis (Figure 1). Considering they are formed from two subunits of non-equal size, they are slightly longer in the centrality than in diameter.
Prokaryotic ribosomes [edit]
Prokaryotic ribosomes are around twenty nm (200 Å) in diameter and are composed of 65% rRNA and 35% ribosomal proteins.[12] Eukaryotic ribosomes are between 25 and 30 nm (250–300 Å) in diameter with an rRNA-to-protein ratio that is close to 1.[13] Crystallographic piece of work[14] has shown that there are no ribosomal proteins close to the reaction site for polypeptide synthesis. This suggests that the poly peptide components of ribosomes do not straight participate in peptide bond formation catalysis, just rather that these proteins human action as a scaffold that may heighten the power of rRNA to synthesize protein (See: Ribozyme).
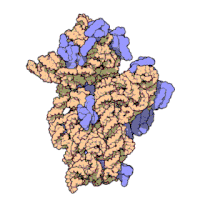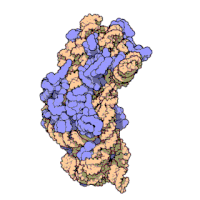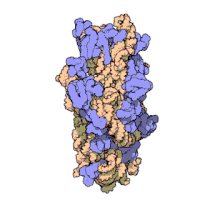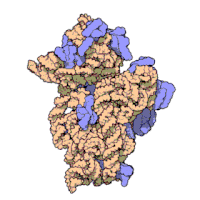
Effigy three: Molecular structure of the 30S subunit from Thermus thermophilus.[xv] Proteins are shown in blueish and the single RNA concatenation in brown.
The ribosomal subunits of prokaryotes and eukaryotes are quite similar.[xvi]
The unit of measurement used to describe the ribosomal subunits and the rRNA fragments is the Svedberg unit of measurement, a measure of the rate of sedimentation in centrifugation rather than size. This accounts for why fragment names do not add up: for case, bacterial 70S ribosomes are made of 50S and 30S subunits.
Prokaryotes take 70S ribosomes, each consisting of a small (30S) and a large (50S) subunit. E. coli, for case, has a 16S RNA subunit (consisting of 1540 nucleotides) that is bound to 21 proteins. The large subunit is composed of a 5S RNA subunit (120 nucleotides), a 23S RNA subunit (2900 nucleotides) and 31 proteins.[16]
-
Ribosome of Due east. coli (a bacterium)[17] : 962 ribosome subunit rRNAs r-proteins 70S 50S 23S (2904 nt) 31 5S (120 nt) 30S 16S (1542 nt) 21
Affinity label for the tRNA binding sites on the Eastward. coli ribosome allowed the identification of A and P site proteins most likely associated with the peptidyltransferase activity;[5] labelled proteins are L27, L14, L15, L16, L2; at least L27 is located at the donor site, as shown by East. Collatz and A.P. Czernilofsky.[18] [nineteen] Additional research has demonstrated that the S1 and S21 proteins, in association with the iii′-end of 16S ribosomal RNA, are involved in the initiation of translation.[20]
Archaeal ribosomes [edit]
Archaeal ribosomes share the same general dimensions of bacteria ones, beingness a 70S ribosome fabricated up from a 50S large subunit, a 30S minor subunit, and containing three rRNA chains. However, on the sequence level, they are much closer to eukaryotic ones than to bacterial ones. Every extra ribosomal protein archaea accept compared to bacteria has a eukaryotic counterpart, while no such relation applies between archaea and bacteria.[21] [22] [23]
Eukaryotic ribosomes [edit]
Eukaryotes have 80S ribosomes located in their cytosol, each consisting of a minor (40S) and large (60S) subunit. Their 40S subunit has an 18S RNA (1900 nucleotides) and 33 proteins.[24] [25] The large subunit is equanimous of a 5S RNA (120 nucleotides), 28S RNA (4700 nucleotides), a 5.8S RNA (160 nucleotides) subunits and 46 proteins.[16] [24] [26]
-
eukaryotic cytosolic ribosomes (R. norvegicus)[17] : 65 ribosome subunit rRNAs r-proteins 80S 60S 28S (4718 nt) 49 5.8S (160 nt) 5S (120 nt) 40S 18S (1874 nt) 33
During 1977, Czernilofsky published enquiry that used affinity labeling to identify tRNA-binding sites on rat liver ribosomes. Several proteins, including L32/33, L36, L21, L23, L28/29 and L13 were implicated as beingness at or near the peptidyl transferase heart.[27]
Plastoribosomes and mitoribosomes [edit]
In eukaryotes, ribosomes are nowadays in mitochondria (sometimes called mitoribosomes) and in plastids such as chloroplasts (besides called plastoribosomes). They besides consist of large and minor subunits bound together with proteins into one 70S particle.[16] These ribosomes are similar to those of leaner and these organelles are idea to have originated as symbiotic leaner[16] Of the two, chloroplastic ribosomes are closer to bacterial ones than mitochrondrial ones are. Many pieces of ribosomal RNA in the mitochrondria are shortened, and in the case of 5S rRNA, replaced by other structures in animals and fungi.[28] In particular, Leishmania tarentolae has a minimalized set up of mitochondrial rRNA.[29] In contrast, establish mitoribosomes have both extended rRNA and additional proteins as compared to bacteria, in item, many pentatricopetide echo proteins.[thirty]
The cryptomonad and chlorarachniophyte algae may contain a nucleomorph that resembles a vestigial eukaryotic nucleus.[31] Eukaryotic 80S ribosomes may be present in the compartment containing the nucleomorph.[32]
Making use of the differences [edit]
The differences between the bacterial and eukaryotic ribosomes are exploited by pharmaceutical chemists to create antibiotics that can destroy a bacterial infection without harming the cells of the infected person. Due to the differences in their structures, the bacterial 70S ribosomes are vulnerable to these antibiotics while the eukaryotic 80S ribosomes are not.[33] Even though mitochondria possess ribosomes similar to the bacterial ones, mitochondria are not affected by these antibiotics considering they are surrounded past a double membrane that does not easily admit these antibiotics into the organelle.[34] A noteworthy counterexample, notwithstanding, includes the antineoplastic antibiotic chloramphenicol, which successfully inhibits bacterial 50S and eukaryotic mitochondrial 50S ribosomes.[35] The aforementioned of mitochondria cannot be said of chloroplasts, where antibiotic resistance in ribosomal proteins is a trait to be introduced as a mark in genetic engineering.[36]
Common properties [edit]
The diverse ribosomes share a core structure, which is quite similar despite the large differences in size. Much of the RNA is highly organized into diverse 3rd structural motifs, for example pseudoknots that exhibit coaxial stacking. The extra RNA in the larger ribosomes is in several long continuous insertions,[37] such that they form loops out of the core structure without disrupting or irresolute information technology.[16] All of the catalytic activity of the ribosome is carried out past the RNA; the proteins reside on the surface and seem to stabilize the structure.[sixteen]
High-resolution structure [edit]

Figure four: Atomic structure of the 50S subunit from Haloarcula marismortui. Proteins are shown in blue and the two RNA bondage in brown and yellow.[38] The small patch of dark-green in the center of the subunit is the active site.
The full general molecular structure of the ribosome has been known since the early on 1970s. In the early on 2000s, the structure has been accomplished at high resolutions, of the order of a few ångströms.
The kickoff papers giving the structure of the ribosome at atomic resolution were published almost simultaneously in late 2000. The 50S (large prokaryotic) subunit was adamant from the archaeon Haloarcula marismortui [38] and the bacterium Deinococcus radiodurans,[39] and the structure of the 30S subunit was determined from Thermus thermophilus.[15] These structural studies were awarded the Nobel Prize in Chemical science in 2009. In May 2001 these coordinates were used to reconstruct the entire T. thermophilus 70S particle at v.5 Å resolution.[40]
Two papers were published in Nov 2005 with structures of the Escherichia coli 70S ribosome. The structures of a vacant ribosome were determined at iii.5 Å resolution using X-ray crystallography.[41] And so, two weeks afterward, a structure based on cryo-electron microscopy was published,[42] which depicts the ribosome at 11–15 Å resolution in the human activity of passing a newly synthesized poly peptide strand into the protein-conducting channel.
The first atomic structures of the ribosome complexed with tRNA and mRNA molecules were solved by using X-ray crystallography by two groups independently, at 2.viii Å[43] and at 3.vii Å.[44] These structures allow one to come across the details of interactions of the Thermus thermophilus ribosome with mRNA and with tRNAs spring at classical ribosomal sites. Interactions of the ribosome with long mRNAs containing Shine-Dalgarno sequences were visualized soon after that at 4.5–v.five Å resolution.[45]
In 2011, the starting time consummate diminutive structure of the eukaryotic 80S ribosome from the yeast Saccharomyces cerevisiae was obtained by crystallography.[24] The model reveals the compages of eukaryote-specific elements and their interaction with the universally conserved core. At the same time, the complete model of a eukaryotic 40S ribosomal structure in Tetrahymena thermophila was published and described the structure of the 40S subunit, as well every bit much about the 40S subunit'due south interaction with eIF1 during translation initiation.[25] Similarly, the eukaryotic 60S subunit structure was too determined from Tetrahymena thermophila in circuitous with eIF6.[26]
Office [edit]
Ribosomes are minute particles consisting of RNA and associated proteins that function to synthesize proteins. Proteins are needed for many cellular functions such as repairing impairment or directing chemical processes. Ribosomes tin exist found floating inside the cytoplasm or attached to the endoplasmic reticulum. Their main function is to catechumen genetic lawmaking into an amino acid sequence and to build protein polymers from amino acid monomers.
Ribosomes act equally catalysts in two extremely important biological processes chosen peptidyl transfer and peptidyl hydrolysis [v] [46] The "PT heart is responsible for producing poly peptide bonds during protein elongation".[46]
In summary, ribosomes accept two main functions: decoding the message and the germination of peptide bonds. These two functions reside in the ribosomal subunits. Each subunit is made of one or more than rRNAs and many r-proteins. The small subunit (30S in leaner and archaea, 40S in eukaryotes) has the decoding function, whereas the large subunit (50S in bacteria and archaea, 60S in eukaryotes) catalyzes the germination of peptide bonds, referred to every bit the peptidyl-transferase activity. The bacterial (and archaeal) small subunit contains the 16S rRNA and 21 r-proteins (Escherichia coli), whereas the eukaryotic pocket-sized subunit contains the 18S rRNA and 32 r-proteins (Saccharomyces cerevisiae; although the numbers vary between species). The bacterial large subunit contains the 5S and 23S rRNAs and 34 r-proteins (E. coli), with the eukaryotic large subunit containing the 5S, five.8S and 25S/28S rRNAs and 46 r-proteins (Due south. cerevisiae; again, the exact numbers vary betwixt species).[47]
Translation [edit]
Ribosomes are the workplaces of protein biosynthesis, the process of translating mRNA into poly peptide. The mRNA comprises a series of codons which are decoded by the ribosome so equally to brand the protein. Using the mRNA equally a template, the ribosome traverses each codon (3 nucleotides) of the mRNA, pairing it with the appropriate amino acid provided by an aminoacyl-tRNA. Aminoacyl-tRNA contains a complementary anticodon on one end and the advisable amino acid on the other. For fast and accurate recognition of the advisable tRNA, the ribosome utilizes large conformational changes (conformational proofreading).[48] The pocket-size ribosomal subunit, typically bound to an aminoacyl-tRNA containing the get-go amino acid methionine, binds to an AUG codon on the mRNA and recruits the large ribosomal subunit. The ribosome contains iii RNA binding sites, designated A, P and E. The A-site binds an aminoacyl-tRNA or termination release factors;[49] [l] the P-site binds a peptidyl-tRNA (a tRNA bound to the poly-peptide concatenation); and the E-site (exit) binds a costless tRNA. Poly peptide synthesis begins at a start codon AUG near the 5' end of the mRNA. mRNA binds to the P site of the ribosome first. The ribosome recognizes the start codon by using the Shine-Dalgarno sequence of the mRNA in prokaryotes and Kozak box in eukaryotes.
Although catalysis of the peptide bond involves the C2 hydroxyl of RNA'south P-site adenosine in a proton shuttle mechanism, other steps in protein synthesis (such as translocation) are acquired past changes in poly peptide conformations. Since their catalytic core is made of RNA, ribosomes are classified as "ribozymes,"[51] and information technology is thought that they might be remnants of the RNA world.[52]

Figure 5: Translation of mRNA (one) by a ribosome (2)(shown as small and big subunits) into a polypeptide concatenation (3). The ribosome begins at the get-go codon of RNA (AUG) and ends at the terminate codon (UAG).
In Figure 5, both ribosomal subunits (small-scale and large) assemble at the start codon (towards the 5' cease of the mRNA). The ribosome uses tRNA that matches the current codon (triplet) on the mRNA to suspend an amino acid to the polypeptide chain. This is done for each triplet on the mRNA, while the ribosome moves towards the three' terminate of the mRNA. Usually in bacterial cells, several ribosomes are working parallel on a unmarried mRNA, forming what is called a polyribosome or polysome.
Cotranslational folding [edit]
The ribosome is known to actively participate in the protein folding.[53] [54] The structures obtained in this way are usually identical to the ones obtained during protein chemical refolding; still, the pathways leading to the final product may be dissimilar.[55] [56] In some cases, the ribosome is crucial in obtaining the functional protein course. For example, 1 of the possible mechanisms of folding of the deeply knotted proteins relies on the ribosome pushing the chain through the attached loop.[57]
Addition of translation-contained amino acids [edit]
Presence of a ribosome quality control protein Rqc2 is associated with mRNA-contained protein elongation.[58] [59] This elongation is a result of ribosomal addition (via tRNAs brought past Rqc2) of Cat tails: ribosomes extend the C -terminus of a stalled protein with random, translation-independent sequences of a lanines and t hreonines.[sixty] [61]
Ribosome locations [edit]
Ribosomes are classified as being either "free" or "membrane-bound".
![]()
Free and membrane-leap ribosomes differ merely in their spatial distribution; they are identical in structure. Whether the ribosome exists in a gratis or membrane-spring state depends on the presence of an ER-targeting signal sequence on the protein being synthesized, and so an individual ribosome might be membrane-bound when it is making one protein, but complimentary in the cytosol when information technology makes another protein.
Ribosomes are sometimes referred to as organelles, but the employ of the term organelle is often restricted to describing sub-cellular components that include a phospholipid membrane, which ribosomes, being entirely particulate, do non. For this reason, ribosomes may sometimes be described as "not-membranous organelles".
Free ribosomes [edit]
Free ribosomes can move nigh anywhere in the cytosol, but are excluded from the cell nucleus and other organelles. Proteins that are formed from costless ribosomes are released into the cytosol and used within the prison cell. Since the cytosol contains high concentrations of glutathione and is, therefore, a reducing environs, proteins containing disulfide bonds, which are formed from oxidized cysteine residues, cannot be produced within information technology.
Membrane-leap ribosomes [edit]
When a ribosome begins to synthesize proteins that are needed in some organelles, the ribosome making this protein can become "membrane-bound". In eukaryotic cells this happens in a region of the endoplasmic reticulum (ER) called the "crude ER". The newly produced polypeptide chains are inserted direct into the ER by the ribosome undertaking vectorial synthesis and are so transported to their destinations, through the secretory pathway. Bound ribosomes usually produce proteins that are used within the plasma membrane or are expelled from the cell via exocytosis.[62]
Biogenesis [edit]
In bacterial cells, ribosomes are synthesized in the cytoplasm through the transcription of multiple ribosome gene operons. In eukaryotes, the process takes identify both in the jail cell cytoplasm and in the nucleolus, which is a region inside the cell nucleus. The assembly process involves the coordinated function of over 200 proteins in the synthesis and processing of the four rRNAs, likewise every bit associates of those rRNAs with the ribosomal proteins.
Origin [edit]
The ribosome may have first originated in an RNA world, appearing as a cocky-replicating complex that only later evolved the ability to synthesize proteins when amino acids began to appear.[63] Studies propose that ancient ribosomes constructed solely of rRNA could take developed the ability to synthesize peptide bonds.[64] [65] [66] In addition, evidence strongly points to ancient ribosomes every bit cocky-replicating complexes, where the rRNA in the ribosomes had informational, structural, and catalytic purposes considering information technology could have coded for tRNAs and proteins needed for ribosomal cocky-replication.[67] Hypothetical cellular organisms with self-replicating RNA but without DNA are called ribocytes (or ribocells).[68] [69]
Equally amino acids gradually appeared in the RNA earth under prebiotic conditions,[70] [71] their interactions with catalytic RNA would increment both the range and efficiency of office of catalytic RNA molecules.[63] Thus, the driving force for the evolution of the ribosome from an ancient self-replicating machine into its current form every bit a translational automobile may have been the selective pressure level to incorporate proteins into the ribosome's self-replicating mechanisms, so as to increment its capacity for cocky-replication.[67] [72] [73]
Heterogeneous ribosomes [edit]
Ribosomes are compositionally heterogeneous between species and fifty-fifty inside the same jail cell, as evidenced by the beingness of cytoplasmic and mitochondria ribosomes within the aforementioned eukaryotic cells. Certain researchers accept suggested that heterogeneity in the composition of ribosomal proteins in mammals is important for factor regulation, i.e., the specialized ribosome hypothesis.[74] [75] However, this hypothesis is controversial and the topic of ongoing inquiry.[76] [77]
Heterogeneity in ribosome limerick was offset proposed to be involved in translational control of protein synthesis by Vince Mauro and Gerald Edelman.[78] They proposed the ribosome filter hypothesis to explain the regulatory functions of ribosomes. Bear witness has suggested that specialized ribosomes specific to different cell populations may affect how genes are translated.[79] Some ribosomal proteins exchange from the assembled complex with cytosolic copies[80] suggesting that the structure of the in vivo ribosome can be modified without synthesizing an entire new ribosome.
Certain ribosomal proteins are admittedly critical for cellular life while others are not. In budding yeast, 14/78 ribosomal proteins are non-essential for growth, while in humans this depends on the cell of written report.[81] Other forms of heterogeneity include post-translational modifications to ribosomal proteins such every bit acetylation, methylation, and phosphorylation.[82] Arabidopsis,[83] [84] [85] [86] Viral internal ribosome entry sites (IRESs) may mediate translations by compositionally distinct ribosomes. For instance, 40S ribosomal units without eS25 in yeast and mammalian cells are unable to recruit the CrPV IGR IRES.[87]
Heterogeneity of ribosomal RNA modifications plays an important role in structural maintenance and/or function and almost mRNA modifications are found in highly conserved regions.[88] [89] The nearly common rRNA modifications are pseudouridylation and 2'-O methylation of ribose.[xc]
See also [edit]
- Aminoglycosides
- Biological machines
- Posttranslational modification
- Protein dynamics
- RNA 3rd structure
- Translation (genetics)
- Wobble base pair
- Ada Yonath—Israeli crystallographer known for her pioneering piece of work on the structure of the ribosome, for which she won the Nobel Prize.
References [edit]
- ^ Konikkat S (February 2016). Dynamic Remodeling Events Drive the Removal of the ITS2 Spacer Sequence During Associates of 60S Ribosomal Subunits in South. cerevisiae (Ph.D. thesis). Carnegie Mellon University. Archived from the original on 3 August 2017.
- ^ Weiler EW, Nover 50 (2008). Allgemeine und Molekulare Botanik (in German). Stuttgart: Georg Thieme Verlag. p. 532. ISBN978-3-13-152791-2.
- ^ de la Cruz J, Karbstein G, Woolford JL (2015). "Functions of ribosomal proteins in associates of eukaryotic ribosomes in vivo". Annual Review of Biochemistry. 84: 93–129. doi:10.1146/annurev-biochem-060614-033917. PMC4772166. PMID 25706898.
- ^ "Scitable past nature translation / RNA translation".
- ^ a b c Tirumalai MR, Rivas M, Tran Q, Fox GE (November 2021). "The Peptidyl Transferase Center: a Window to the By". Microbiol Mol Biol Rev. 85 (four): e0010421. doi:10.1128/MMBR.00104-21. PMC 8579967. PMID 34756086.
- ^ Benne R, Sloof P (1987). "Evolution of the mitochondrial protein synthetic machinery". Bio Systems. 21 (1): 51–68. doi:10.1016/0303-2647(87)90006-2. PMID 2446672.
- ^ "Ribosomes". Archived from the original on 2009-03-xx. Retrieved 2011-04-28 .
- ^ Palade GE (January 1955). "A small particulate component of the cytoplasm". The Journal of Biophysical and Biochemical Cytology. 1 (1): 59–68. doi:10.1083/jcb.1.1.59. PMC2223592. PMID 14381428.
- ^ Roberts RB, ed. (1958). "Introduction". Microsomal Particles and Protein Synthesis. New York: Pergamon Press, Inc.
- ^ "The Nobel Prize in Physiology or Medicine 1974". Nobelprize.org. The Nobel Foundation. Archived from the original on 26 Jan 2013. Retrieved 10 December 2012.
- ^ "2009 Nobel Prize in Chemistry". The Nobel Foundation. Archived from the original on 28 April 2012. Retrieved 10 December 2012.
- ^ Kurland CG (1960). "Molecular characterization of ribonucleic acid from Escherichia coli ribosomes". Journal of Molecular Biological science. two (two): 83–91. doi:10.1016/s0022-2836(sixty)80029-0.
- ^ Wilson DN, Doudna Cate JH (May 2012). "The structure and office of the eukaryotic ribosome". Cold Bound Harbor Perspectives in Biology. 4 (v): a011536. doi:10.1101/cshperspect.a011536. PMC3331703. PMID 22550233.
- ^ Nissen P, Hansen J, Ban N, Moore Lead, Steitz TA (August 2000). "The structural basis of ribosome activity in peptide bond synthesis" (PDF). Science. 289 (5481): 920–30. Bibcode:2000Sci...289..920N. doi:10.1126/science.289.5481.920. PMID 10937990. S2CID 8370119. Archived from the original (PDF) on 2020-11-30.
- ^ a b Wimberly BT, Brodersen DE, Clemons WM, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan Five (September 2000). "Structure of the 30S ribosomal subunit". Nature. 407 (6802): 327–39. Bibcode:2000Natur.407..327W. doi:10.1038/35030006. PMID 11014182. S2CID 4419944.
- ^ a b c d e f one thousand Alberts B, Johnson A, Lewis J, Raff G, Roberts K, Walter P (2002). "Membrane-bound Ribosomes Define the Crude ER". Molecular Biology of the Cell (4th ed.). New York: Garland Science. p. 342. ISBN978-0-8153-4072-0.
- ^ a b Garrett R, Grisham CM (2009). Biochemistry (4th ed.). Cengage Learning Services. ISBN978-0-495-11464-2.
- ^ Collatz East, Küchler East, Stöffler G, Czernilofsky AP (April 1976). "The site of reaction on ribosomal poly peptide L27 with an analogousness characterization derivative of tRNA Met f". FEBS Letters. 63 (2): 283–half dozen. doi:ten.1016/0014-5793(76)80112-3. PMID 770196.
- ^ Czernilofsky AP, Collatz EE, Stöffler G, Kuechler East (January 1974). "Proteins at the tRNA bounden sites of Escherichia coli ribosomes". Proceedings of the National University of Sciences of the U.s. of America. 71 (one): 230–4. Bibcode:1974PNAS...71..230C. doi:x.1073/pnas.71.one.230. PMC387971. PMID 4589893.
- ^ Czernilofsky AP, Kurland CG, Stöffler G (Oct 1975). "30S ribosomal proteins associated with the 3'-terminus of 16S RNA". FEBS Messages. 58 (1): 281–four. doi:10.1016/0014-5793(75)80279-1. PMID 1225593.
- ^ Cullen KE (2009). "Archaeal Ribosomes". Encyclopedia of Life Science. New York: Facts On File. pp. 1–5. doi:10.1002/9780470015902.a0000293.pub3. ISBN9780470015902. S2CID 243730576.
- ^ Tirumalai MR, Anane-Bediakoh D, Rajesh R, Fox GE (November 2021). "Internet Charges of the Ribosomal Proteins of the S10 and spc Clusters of Halophiles Are Inversely Related to the Degree of Halotolerance". Microbiol. Spectrum. 9 (iii): e0178221. doi:10.1128/spectrum.01782-21. PMC8672879. PMID 34908470.
- ^ Wang J, Dasgupta I, Fob GE (28 Apr 2009). "Many nonuniversal archaeal ribosomal proteins are found in conserved gene clusters". Archaea. ii (four): 241–51. doi:x.1155/2009/971494. PMC2686390. PMID 19478915.
- ^ a b c Ben-Shem A, Garreau de Loubresse N, Melnikov S, Jenner L, Yusupova G, Yusupov K (December 2011). "The structure of the eukaryotic ribosome at iii.0 Å resolution". Science. 334 (6062): 1524–9. Bibcode:2011Sci...334.1524B. doi:10.1126/scientific discipline.1212642. PMID 22096102. S2CID 9099683.
- ^ a b Rabl J, Leibundgut M, Ataide SF, Haag A, Ban N (February 2011). "Crystal structure of the eukaryotic 40S ribosomal subunit in complex with initiation factor 1" (PDF). Science. 331 (6018): 730–6. Bibcode:2011Sci...331..730R. doi:x.1126/science.1198308. hdl:20.500.11850/153130. PMID 21205638. S2CID 24771575.
- ^ a b Klinge S, Voigts-Hoffmann F, Leibundgut Thousand, Arpagaus S, Ban N (November 2011). "Crystal construction of the eukaryotic 60S ribosomal subunit in complex with initiation factor half dozen". Scientific discipline. 334 (6058): 941–8. Bibcode:2011Sci...334..941K. doi:10.1126/science.1211204. PMID 22052974. S2CID 206536444.
- ^ Fabijanski Southward, Pellegrini G (1977). "Identification of proteins at the peptidyl-tRNA bounden site of rat liver ribosomes". Molecular & Full general Genetics. 184 (3): 551–vi. doi:10.1007/BF00431588. PMID 6950200. S2CID 9751945.
- ^ Agrawal RK, Sharma MR (December 2012). "Structural aspects of mitochondrial translational apparatus". Electric current Stance in Structural Biology. 22 (vi): 797–803. doi:10.1016/j.sbi.2012.08.003. PMC3513651. PMID 22959417.
- ^ Sharma MR, Booth TM, Simpson L, Maslov DA, Agrawal RK (June 2009). "Structure of a mitochondrial ribosome with minimal RNA". Proceedings of the National Academy of Sciences of the United states of america of America. 106 (24): 9637–42. Bibcode:2009PNAS..106.9637S. doi:10.1073/pnas.0901631106. PMC2700991. PMID 19497863.
- ^ Waltz F, Nguyen TT, Arrivé Thousand, Bochler A, Chicher J, Hammann P, Kuhn L, Quadrado M, Mireau H, Hashem Y, Giegé P (Jan 2019). "Small is big in Arabidopsis mitochondrial ribosome". Nature Plants. 5 (1): 106–117. doi:10.1038/s41477-018-0339-y. PMID 30626926. S2CID 58004990.
- ^ Archibald JM, Lane CE (2009). "Going, going, not quite gone: nucleomorphs as a case report in nuclear genome reduction". The Journal of Heredity. 100 (five): 582–90. doi:ten.1093/jhered/esp055. PMID 19617523.
- ^ "Specialized Internal Structures of Prokaryotes | Boundless Microbiology". courses.lumenlearning.com . Retrieved 2021-09-24 .
- ^ Recht MI, Douthwaite S, Puglisi JD (June 1999). "Footing for prokaryotic specificity of action of aminoglycoside antibiotics". The EMBO Journal. 18 (11): 3133–8. doi:10.1093/emboj/18.11.3133. PMC1171394. PMID 10357824.
- ^ O'Brien TW (May 1971). "The general occurrence of 55 S ribosomes in mammalian liver mitochondria". The Journal of Biological Chemistry. 246 (ten): 3409–17. doi:10.1016/S0021-9258(xviii)62239-2. PMID 4930061.
- ^ "Chloramphenicol-lnduced Bone Marrow Suppression". JAMA. 213 (7): 1183–1184. 1970-08-17. doi:10.1001/jama.1970.03170330063011. ISSN 0098-7484. PMID 5468266.
- ^ Newman SM, Boynton JE, Gillham NW, Randolph-Anderson BL, Johnson AM, Harris EH (Dec 1990). "Transformation of chloroplast ribosomal RNA genes in Chlamydomonas: molecular and genetic characterization of integration events". Genetics. 126 (4): 875–88. doi:x.1093/genetics/126.iv.875. PMC1204285. PMID 1981764.
- ^ Penev PI, Fakhretaha-Aval South, Patel VJ, Cannone JJ, Gutell RR, Petrov Equally, Williams LD, Glass JB (Baronial 2020). "Supersized ribosomal RNA expansion segments in Asgard archaea". Genome Biology and Development. 12 (ten): 1694–1710. doi:ten.1093/gbe/evaa170. PMC7594248. PMID 32785681.
- ^ a b Ban Northward, Nissen P, Hansen J, Moore PB, Steitz TA (August 2000). "The complete atomic structure of the large ribosomal subunit at 2.4 A resolution". Science. 289 (5481): 905–20. Bibcode:2000Sci...289..905B. CiteSeerX10.1.1.58.2271. doi:ten.1126/science.289.5481.905. PMID 10937989.
- ^ Schluenzen F, Tocilj A, Zarivach R, Harms J, Gluehmann M, Janell D, Bashan A, Bartels H, Agmon I, Franceschi F, Yonath A (September 2000). "Structure of functionally activated small ribosomal subunit at three.three angstroms resolution". Prison cell. 102 (5): 615–23. doi:10.1016/S0092-8674(00)00084-ii. PMID 11007480. S2CID 1024446.
- ^ Yusupov MM, Yusupova GZ, Baucom A, Lieberman Grand, Earnest TN, Cate JH, Noller HF (May 2001). "Crystal structure of the ribosome at v.5 A resolution". Science. 292 (5518): 883–96. Bibcode:2001Sci...292..883Y. doi:ten.1126/scientific discipline.1060089. PMID 11283358. S2CID 39505192.
- ^ Schuwirth BS, Borovinskaya MA, Hau CW, Zhang W, Vila-Sanjurjo A, Holton JM, Cate JH (November 2005). "Structures of the bacterial ribosome at 3.5 A resolution". Science. 310 (5749): 827–34. Bibcode:2005Sci...310..827S. doi:10.1126/science.1117230. PMID 16272117. S2CID 37382005.
- ^ Mitra G, Schaffitzel C, Shaikh T, Tama F, Jenni S, Brooks CL, Ban N, Frank J (November 2005). "Structure of the E. coli poly peptide-conducting channel bound to a translating ribosome". Nature. 438 (7066): 318–24. Bibcode:2005Natur.438..318M. doi:10.1038/nature04133. PMC1351281. PMID 16292303.
- ^ Selmer Thousand, Dunham CM, Spud FV, Weixlbaumer A, Petry S, Kelley Ac, Weir JR, Ramakrishnan V (September 2006). "Structure of the 70S ribosome complexed with mRNA and tRNA". Science. 313 (5795): 1935–42. Bibcode:2006Sci...313.1935S. doi:10.1126/science.1131127. PMID 16959973. S2CID 9737925.
- ^ Korostelev A, Trakhanov Due south, Laurberg M, Noller HF (September 2006). "Crystal construction of a 70S ribosome-tRNA complex reveals functional interactions and rearrangements". Cell. 126 (half-dozen): 1065–77. doi:10.1016/j.prison cell.2006.08.032. PMID 16962654. S2CID 13452915.
- ^ Yusupova G, Jenner Fifty, Rees B, Moras D, Yusupov M (Nov 2006). "Structural basis for messenger RNA move on the ribosome". Nature. 444 (7117): 391–4. Bibcode:2006Natur.444..391Y. doi:10.1038/nature05281. PMID 17051149. S2CID 4419198.
- ^ a b "Specialized Internal Structures of Prokaryotes | Boundless Microbiology". courses.lumenlearning.com . Retrieved 2018-09-27 .
- ^ Lafontaine, D., Tollervey, D. The function and synthesis of ribosomes. Nat Rev Mol Cell Biol two, 514–520 (2001). https://doi.org/x.1038/35080045
- ^ Savir Y, Tlusty T (April 2013). "The ribosome equally an optimal decoder: a lesson in molecular recognition". Jail cell. 153 (two): 471–9. Bibcode:2013APS..MARY46006T. doi:10.1016/j.cell.2013.03.032. PMID 23582332.
- ^ Korkmaz M, Sanyal South (September 2017). "Escherichia coli". The Journal of Biological Chemistry. 292 (36): 15134–15142. doi:10.1074/jbc.M117.785238. PMC5592688. PMID 28743745.
- ^ Konevega AL, Soboleva NG, Makhno Vi, Semenkov YP, Wintermeyer Due west, Rodnina MV, Katunin VI (January 2004). "Purine bases at position 37 of tRNA stabilize codon-anticodon interaction in the ribosomal A site by stacking and Mg2+-dependent interactions". RNA. 10 (ane): xc–101. doi:10.1261/rna.5142404. PMC1370521. PMID 14681588.
- ^ Rodnina MV, Beringer 1000, Wintermeyer W (January 2007). "How ribosomes make peptide bonds". Trends in Biochemical Sciences. 32 (ane): 20–6. doi:10.1016/j.tibs.2006.eleven.007. PMID 17157507.
- ^ Cech TR (Baronial 2000). "Structural biology. The ribosome is a ribozyme". Science. 289 (5481): 878–nine. doi:x.1126/scientific discipline.289.5481.878. PMID 10960319. S2CID 24172338.
- ^ Banerjee D, Sanyal S (October 2014). "Poly peptide folding activity of the ribosome (PFAR) -- a target for antiprion compounds". Viruses. 6 (ten): 3907–24. doi:10.3390/v6103907. PMC4213570. PMID 25341659.
- ^ Fedorov AN, Baldwin TO (December 1997). "Cotranslational protein folding". The Journal of Biological Chemistry. 272 (52): 32715–8. doi:10.1074/jbc.272.52.32715. PMID 9407040.
- ^ Baldwin RL (June 1975). "Intermediates in protein folding reactions and the mechanism of protein folding". Annual Review of Biochemistry. 44 (1): 453–75. doi:10.1146/annurev.bi.44.070175.002321. PMID 1094916.
- ^ Das D, Das A, Samanta D, Ghosh J, Dasgupta Due south, Bhattacharya A, Basu A, Sanyal S, Das Gupta C (Baronial 2008). "Part of the ribosome in protein folding" (PDF). Biotechnology Journal. 3 (8): 999–1009. doi:10.1002/biot.200800098. PMID 18702035.
- ^ Dabrowski-Tumanski P, Piejko M, Niewieczerzal Southward, Stasiak A, Sulkowska JI (December 2018). "Protein Knotting by Agile Threading of Nascent Polypeptide Chain Exiting from the Ribosome Exit Aqueduct". The Periodical of Physical Chemistry B. 122 (49): 11616–11625. doi:10.1021/acs.jpcb.8b07634. PMID 30198720. S2CID 52176392.
- ^ Brandman O, Stewart-Ornstein J, Wong D, Larson A, Williams CC, Li GW, Zhou South, Rex D, Shen PS, Weibezahn J, Dunn JG, Rouskin S, Inada T, Frost A, Weissman JS (November 2012). "A ribosome-bound quality control complex triggers deposition of nascent peptides and signals translation stress". Jail cell. 151 (5): 1042–54. doi:10.1016/j.cell.2012.x.044. PMC3534965. PMID 23178123.
- ^ Defenouillère Q, Yao Y, Mouaikel J, Namane A, Galopier A, Decourty Fifty, Doyen A, Malabat C, Saveanu C, Jacquier A, Fromont-Racine M (March 2013). "Cdc48-associated complex bound to 60S particles is required for the clearance of aberrant translation products". Proceedings of the National Academy of Sciences of the United states of america of America. 110 (13): 5046–51. Bibcode:2013PNAS..110.5046D. doi:ten.1073/pnas.1221724110. PMC3612664. PMID 23479637.
- ^ Shen PS, Park J, Qin Y, Li X, Parsawar K, Larson MH, Cox J, Cheng Y, Lambowitz AM, Weissman JS, Brandman O, Frost A (January 2015). "Poly peptide synthesis. Rqc2p and 60S ribosomal subunits mediate mRNA-contained elongation of nascent chains". Science. 347 (6217): 75–8. Bibcode:2015Sci...347...75S. doi:10.1126/science.1259724. PMC4451101. PMID 25554787.
- ^ Keeley J, Gutnikoff R (2015-01-02). "Ribosome Studies Plough Upward New Mechanism of Protein Synthesis" (Press release). Howard Hughes Medical Institute. Archived from the original on 2015-01-12. Retrieved 2015-01-16 .
- ^ Alberts B, Johnson A, Lewis J, Raff Chiliad, Roberts Thousand, Walter P (2002). "Membrane-bound Ribosomes Ascertain the Rough ER". Molecular Biology of the Cell (4th ed.). New York: Garland Science. ISBN978-0-8153-4072-0.
- ^ a b Noller HF (Apr 2012). "Evolution of protein synthesis from an RNA world". Cold Spring Harbor Perspectives in Biology. iv (4): a003681. doi:10.1101/cshperspect.a003681. PMC3312679. PMID 20610545.
- ^ Dabbs ER (1986). Mutant studies on the prokaryotic ribosome. New York: Springer-Verlag.
- ^ Noller HF, Hoffarth 5, Zimniak 50 (June 1992). "Unusual resistance of peptidyl transferase to protein extraction procedures". Science. 256 (5062): 1416–nine. Bibcode:1992Sci...256.1416N. doi:10.1126/science.1604315. PMID 1604315.
- ^ Nomura M, Mizushima S, Ozaki M, Traub P, Lowry CV (1969). "Construction and function of ribosomes and their molecular components". Cold Spring Harbor Symposia on Quantitative Biology. 34: 49–61. doi:x.1101/sqb.1969.034.01.009. PMID 4909519.
- ^ a b Root-Bernstein Thou, Root-Bernstein R (February 2015). "The ribosome equally a missing link in the evolution of life". Journal of Theoretical Biological science. 367: 130–158. doi:10.1016/j.jtbi.2014.xi.025. PMID 25500179.
- ^ Yarus M (2002). "Primordial genetics: phenotype of the ribocyte". Annual Review of Genetics. 36: 125–51. doi:10.1146/annurev.genet.36.031902.105056. PMID 12429689.
- ^ Forterre P, Krupovic Thou (2012). "The Origin of Virions and Virocells: The Escape Hypothesis Revisited". Viruses: Essential Agents of Life. pp. 43–60. doi:10.1007/978-94-007-4899-6_3. ISBN978-94-007-4898-ix.
- ^ Caetano-Anollés G, Seufferheld MJ (2013). "The coevolutionary roots of biochemistry and cellular organization challenge the RNA globe paradigm". Journal of Molecular Microbiology and Biotechnology. 23 (one–2): 152–77. doi:10.1159/000346551. PMID 23615203. S2CID 41725226.
- ^ Saladino R, Botta Thou, Pino S, Costanzo G, Di Mauro E (August 2012). "Genetics commencement or metabolism start? The formamide clue". Chemical Society Reviews. 41 (xvi): 5526–65. doi:10.1039/c2cs35066a. PMID 22684046.
- ^ Play a joke on GE (September 2010). "Origin and Evolution of the Ribosome". Cold Spring Harb Perspect Biol. 2 (nine): a003483. doi:10.1101/cshperspect.a003483. PMC2926754. PMID 20534711.
- ^ Fox GE (2016). "Origins and early on evolution of the ribosome". In Hernández G, Jagus R (eds.). Evolution of the Protein Synthesis Mechanism and Its Regulation. Switzerland: Springer, Cham. pp. 31–threescore. doi:10.1007/978-3-319-39468-eight. ISBN978-iii-319-39468-eight. S2CID 27493054.
- ^ Shi Z, Fujii K, Kovary KM, Genuth NR, Röst HL, Teruel MN, Barna M (July 2017). "Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide". Molecular Cell. Elsevier BV. 67 (1): 71–83.e7. doi:x.1016/j.molcel.2017.05.021. PMC5548184. PMID 28625553.
- ^ Xue Southward, Barna M (May 2012). "Specialized ribosomes: a new frontier in gene regulation and organismal biology". Nature Reviews. Molecular Prison cell Biology. Springer Science and Business Media LLC. xiii (6): 355–369. doi:ten.1038/nrm3359. PMC4039366. PMID 22617470.
- ^ Ferretti MB, Karbstein K (May 2019). "Does functional specialization of ribosomes really exist?". RNA. Cold Spring Harbor Laboratory. 25 (5): 521–538. doi:10.1261/rna.069823.118. PMC6467006. PMID 30733326.
- ^ Farley-Barnes KI, Ogawa LM, Baserga SJ (October 2019). "Ribosomopathies: Old Concepts, New Controversies". Trends in Genetics. Elsevier BV. 35 (ten): 754–767. doi:10.1016/j.tig.2019.07.004. PMC6852887. PMID 31376929.
- ^ Mauro VP, Edelman GM (September 2002). "The ribosome filter hypothesis". Proceedings of the National Academy of Sciences of the United states of america. 99 (19): 12031–6. Bibcode:2002PNAS...9912031M. doi:10.1073/pnas.192442499. PMC129393. PMID 12221294.
- ^ Xue S, Barna G (May 2012). "Specialized ribosomes: a new borderland in factor regulation and organismal biological science". Nature Reviews. Molecular Prison cell Biological science. thirteen (6): 355–69. doi:10.1038/nrm3359. PMC4039366. PMID 22617470.
- ^ Mathis AD, Naylor BC, Carson RH, Evans E, Harwell J, Knecht J, Hexem Eastward, Peelor FF, Miller BF, Hamilton KL, Transtrum MK, Bikman BT, Price JC (February 2017). "Mechanisms of In Vivo Ribosome Maintenance Modify in Response to Food Signals". Molecular & Cellular Proteomics. 16 (2): 243–254. doi:x.1074/mcp.M116.063255. PMC5294211. PMID 27932527.
- ^ Steffen KK, McCormick MA, Pham KM, MacKay VL, Delaney JR, Murakami CJ, et al. (May 2012). "Ribosome deficiency protects against ER stress in Saccharomyces cerevisiae". Genetics. Genetics Lodge of America. 191 (i): 107–118. doi:x.1534/genetics.111.136549. PMC3338253. PMID 22377630.
- ^ Lee SW, Berger SJ, Martinović S, Pasa-Tolić L, Anderson GA, Shen Y, et al. (April 2002). "Direct mass spectrometric assay of intact proteins of the yeast big ribosomal subunit using capillary LC/FTICR". Proceedings of the National University of Sciences of the Us of America. 99 (ix): 5942–5947. Bibcode:2002PNAS...99.5942L. doi:10.1073/pnas.082119899. PMC122881. PMID 11983894.
- ^ Carroll AJ, Heazlewood JL, Ito J, Millar AH (Feb 2008). "Analysis of the Arabidopsis cytosolic ribosome proteome provides detailed insights into its components and their mail-translational modification". Molecular & Cellular Proteomics. 7 (2): 347–369. doi:10.1074/mcp.m700052-mcp200. PMID 17934214.
- ^ Odintsova TI, Müller EC, Ivanov AV, Egorov TA, Bienert R, Vladimirov SN, et al. (April 2003). "Label and analysis of posttranslational modifications of the human large cytoplasmic ribosomal subunit proteins by mass spectrometry and Edman sequencing". Journal of Protein Chemical science. 22 (iii): 249–258. doi:x.1023/a:1025068419698. PMID 12962325. S2CID 10710245.
- ^ Yu Y, Ji H, Doudna JA, Leary JA (June 2005). "Mass spectrometric analysis of the human 40S ribosomal subunit: native and HCV IRES-bound complexes". Protein Science. xiv (6): 1438–1446. doi:10.1110/ps.041293005. PMC2253395. PMID 15883184.
- ^ Zeidan Q, Wang Z, De Maio A, Hart GW (June 2010). "O-GlcNAc cycling enzymes associate with the translational machinery and change cadre ribosomal proteins". Molecular Biology of the Cell. 21 (12): 1922–1936. doi:ten.1091/mbc.e09-eleven-0941. PMC2883937. PMID 20410138.
- ^ Landry DM, Hertz MI, Thompson SR (Dec 2009). "RPS25 is essential for translation initiation by the Dicistroviridae and hepatitis C viral IRESs". Genes & Development. 23 (23): 2753–2764. doi:ten.1101/gad.1832209. PMC2788332. PMID 19952110.
- ^ Decatur WA, Fournier MJ (July 2002). "rRNA modifications and ribosome function". Trends in Biochemical Sciences. 27 (7): 344–51. doi:ten.1016/s0968-0004(02)02109-half dozen. PMID 12114023.
- ^ Natchiar SK, Myasnikov AG, Kratzat H, Hazemann I, Klaholz BP (November 2017). "Visualization of chemical modifications in the human 80S ribosome structure". Nature. 551 (7681): 472–477. Bibcode:2017Natur.551..472N. doi:10.1038/nature24482. PMID 29143818. S2CID 4465175.
- ^ Guo H (Baronial 2018). "Specialized ribosomes and the control of translation". Biochemical Society Transactions. 46 (4): 855–869. doi:10.1042/BST20160426. PMID 29986937. S2CID 51609077.
External links [edit]
| | Wikimedia Commons has media related to Ribosomes. |
- Lab computer simulates ribosome in motion
- Role of the Ribosome, Gwen 5. Childs, copied here
- Ribosome in Proteopedia—The gratis, collaborative 3D encyclopedia of proteins & other molecules
- Ribosomal proteins families in ExPASy Archived 2011-04-xxx at the Wayback Auto
- Molecule of the Calendar month Archived 2009-ten-27 at the Wayback Motorcar © RCSB Protein Data Bank:
- Ribosome
- Elongation Factors Archived 2011-03-16 at the Wayback Machine
- Palade
- 3D electron microscopy structures of ribosomes at the EM Data Bank (EMDB)
-
 This commodity incorporates public domain material from the NCBI certificate: "Scientific discipline Primer".
This commodity incorporates public domain material from the NCBI certificate: "Scientific discipline Primer".
Source: https://en.wikipedia.org/wiki/Ribosome
Posted by: zanderspronful1972.blogspot.com
0 Response to "What Do Ribosomes Do In An Animal Cell"
Post a Comment